Efecto de la nutrición sobre la conducta y las emociones
1.Introducción
2. Teorías que relacionan la conducta de ingesta y las emociones
2.1. Teoría psicosomática de Kaplan
2.2. Teoría de Bruch
2.3. Teoría de la obesidad externa/interna de Schachter
2.4. Hipótesis de la restricción
3. Emociones y la conducta de ingesta
3.1. Expectativas y memoria
4. Estrés y la conducta de ingesta
5. Ingesta y estado de ánimo
6.Hormonas implicadas en la ingesta y emociones
7. Epigenética
8. Ingesta emocional
8.1 La ingesta emocional durante la adolescencia
1. Introducción
La metáfora “cajón de sastre” podría definir las aportaciones de la Psicología al ámbito de estudio de las emociones. La disparidad de términos, expresiones, así como leyes y conceptos, hacen que el seguimiento terminológico sea complejo. Expresiones clásicas como emoción y sentimiento, se unen a otras como afecto, estado emocional, estado de ánimo, así como sentimiento positivo, negativo, que se combinan con sentimientos básicos como felicidad, alegría, asco o sorpresa son sólo algunos. Si bien no es objeto de estudio del presente capítulo definir cada uno de ellos, sí serán considerados en relación a la conducta de ingesta.
Los intentos de integración conceptual han sido reiterados en el tiempo. La identificación de seis expresiones faciales universales (Ekman, 1972) dieron lugar a otras tantas emociones básicas: felicidad, ira, miedo, tristeza, repugnancia y sorpresa.
Por otro lado, un accidente fortuito sucedido a mediados del siglo XIX, por un obrero del ferrocarril amplió el estudio del sustrato fisiológico de las emociones. Como es sabido, el cráneo de Phineas Gage fue atravesado por una barrena después de una explosión, sobreviviendo a ella, le produjo cambios radicales en su vida, entre ellas, en el manejo de sus emociones. Durante años el caso fue objeto de estudio, e incluso en este siglo se sigue investigando. Las lesiones sufridas a consecuencia del traumatismo en el lóbulo frontal, en concreto en la corteza prefrontal ventromedial (CPFvm), explicaban la pérdida de capacidad cognitiva y del manejo de las emociones, (Damasio, 1994). Así el paradigma de lesión para comprobar la actividad ha sido el más utilizado hasta que las técnicas de neuroimagen las han ido relegando. De esta forma, diversas investigaciones han podido constatar que las áreas neuroanatómicas implicadas en la emoción humana son la CPFvm, la corteza prefrontal dorsolateral, la corteza cingulada anterior y la ínsula, sin olvidar a la amígdala.
La amígdala está implicada en el condicionamiento del miedo, ampliando la cuestión también a otras áreas de conocimiento de la Psicología, como son la memoria, el aprendizaje y la percepción. Sobre este aspecto son destacables los trabajos sobre la doble vía de procesamiento de las emociones (LeDoux, 1999). Una vía rápida y una lenta conforman éstos, en función de los estímulos visuales percibidos. Aunque esta idea de la doble vía de procesamiento era conocida, LeDoux aporta datos experimentales y conforma la estructura de cada una de ellas.
Las emociones tienen un alto valor para la supervivencia de los sujetos en diferentes contextos y situaciones, al igual que la conducta de ingesta. Así la relación entre la ingesta y las emociones aumenta su valor adaptativo.
2. Teorías que relacionan la conducta de ingesta y las emociones
La suposición de que el afecto y la ingesta están relacionados tiene su origen en la literatura sobre la obesidad. Las teorías anteriores explicaban la sobrealimentación en individuos obesos, mientras que las teorías más recientes apuntan a explicar el comportamiento alimentario en una población de peso normal.
2.1. La teoría psicosomática de Kaplan y Kaplan (1957)
Estos autores propusieron que las personas obesas comen en exceso cuando sienten ansiedad y comer reduce estos valores. El mecanismo por el que se reduce la ansiedad no se entiende completamente, pero puede implicar efectos diferenciales de la ingesta de proteínas y carbohidratos que afecta a la síntesis de neurotransmisores cerebrales, en particular, la serotonina.
Probablemente, también intervienen factores de aprendizaje, como por ejemplo: una asociación anterior con situaciones placenteras y no ansiógenas con la alimentación. Sin embargo, estos autores sintieron que los efectos reductores de la ansiedad de comer no pueden explicarse únicamente sobre la base de los hábitos aprendidos. Plantearon la hipótesis de que existe cierto grado de incompatibilidad fisiológica entre el acto de comer y el miedo o la ansiedad intensa y que, al comer, estas emociones disminuyen temporalmente. Las personas obesas no pueden distinguir entre el hambre y la ansiedad porque aprendieron a comer en respuesta a la ansiedad y al hambre.
2.2. La teoría de Bruch (1973)
Hilde Bruch relacionó la ingesta en exceso con una conciencia defectuosa del hambre. Esta teoría propone que la sensación de hambre no es innata sino aprendida en patrones reconocibles. En el caso de las personas obesas, algo ha fallado en la experiencia y el proceso interpersonal que rodea la satisfacción de las necesidades nutricionales y corporales. Las primeras experiencias incorrectas y confusas habían interferido con su capacidad para reconocer el hambre y la saciedad.
Estas primeras experiencias también habían interferido en la capacidad de diferenciar el hambre, de otras señales de malestar que nada tienen que ver con la privación de alimentos como los estados de tensión emocional suscitados por una variedad de conflictos y problemas. Tales individuos no reconocen cuándo tienen hambre o están saciados, ni diferencian entre la necesidad de comida de otras sensaciones y sentimientos incómodos. Así requieren señales provenientes del exterior para saber cuándo comer y cuánto; ya que su propia conciencia interna no ha sido programada correctamente (Bruch, 1973). Según esta teoría una persona comerá en exceso en respuesta a la tensión emocional y las sensaciones o sentimientos incómodos. Tanto las teorías de Kaplan como las de Bruch llegan a la misma predicción: los individuos obesos comerán en exceso en respuesta a estados emocionales incómodos (Canetti et al., 2002).
2.3. La teoría de la obesidad de interna/externa de Schachter (1968-1971)
Esta teoría hace predicciones diferentes de las teorías descritas anteriormente. Los concomitantes fisiológicos del miedo y de la ansiedad llevarían a las personas del peso normal a suprimir el consumo de alimentos, pero no a las personas con sobrepeso debido a su insensibilidad a las señales internas.
Al igual que Bruch, Schachater planteó la hipótesis de que el reconocimiento de un conjunto de señales fisiológicas, incluídas las contracciones gástricas, interpretadas como hambre, eran un fenómeno aprendido. Las personas con peso normal habían aprendido a etiquetar correctamente las contracciones gástricas como hambre, mientras que las personas con sobrepeso no lo habían hecho. Debido a que las contracciones gástricas disminuyen durante el estrés, las personas con peso normal disminuirían su alimentación cuando se sintieran estresadas, pero tal disminución no tendría ningún efecto sobre la alimentación de los obesos. Como consecuencia de la mala comprensión de las señales fisiológicas internas, las personas obesas confiarían mucho más en las señales externas, tanto para iniciar como dejar de comer. Así la teoría predice que las personas con peso normal podrían disminuir o aumentar su alimentación cuando estuvieran estresadas, mientras que las personas obesas no la disminuirían.
Un primer estudio realizado por Schachter et al. (1968), confirmó esta predicción. Descubrieron que para los sujetos con peso normal, el estrés disminuyó la ingesta entre los que tenían hambre y no tenía efecto en los que no la tenían; mientras que los sujetos con sobrepeso consumieron la misma cantidad de alimentos independientemente de su estado fisiológico. Sin embargo, las investigaciones posteriores no replicaron los hallazgos previos (Lowe y Fisher, 1983; Pine, 1985; Reznick y Balch, 1977; Ruderman, 1983; Slochower et al., 1981), si bien un solo estudio confirmó la predicción, pero solo para alimentos sabrosos (McKenna, 1972).
La cuestión de si las emociones influyen en la conducta alimentaria se ha estudiado a fondo en la población obesa. Aunque los hallazgos están más cerca de las teorías de Kaplan y Bruch en relación a los aspectos psicosomáticos, arrojan luz sobre la complejidad de la conducta alimentaria en las personas obesas.
El impacto del estado de ánimo positivo en la ingesta de alimentos no se ha estudiado tan bien como el de los estados de ánimo negativos. Los estudios que han analizado tales relaciones han arrojado resultados contradictorios. Algunas investigaciones no han encontrado ninguna correlación entre la ingesta de los alimentos y el estado de ánimo positivo (Schmitz, 1996; Davis et al., 1985). Sin embargo, se ha encontrado que la ingesta es mayor tanto para los estados de ánimo positivo como negativos en comparación con un estado de ánimo neutral (Patel y Shlundt, 2001). Estos autores proponen que los efectos del estado de ánimo, ya sean positivos o negativos, implican una desinhibición del control de la alimentación. También sugieren que el estado de ánimo positivo puede aumentar la ingesta de alimentos a través de un mecanismo de aprendizaje asociativo donde la felicidad ha sido asociado con comer más alimentos. Estos hallazgos también pueden estar en la línea de los trabajos de Bruch de que las personas obesas no diferencian la necesidad de alimentos de los estados de tensión emocional, aunque en este caso sea placentera.
Todos los estudios revisados se realizaron en poblaciones obesas y la mayoría de ellos encontraron diferencias significativas entre los sujetos obesos y los grupos de control de peso normal o correlaciones significativas dependientes del peso en el área de alimentación emocional. La conclusión es que las personas obesas se involucran en una alimentación emocional significativamente mayor en comparación con las personas que no son obesas parece bastante sólida y brinda apoyo a las teorías de Kaplan y Bruch en sus aspectos psicosomáticos. Sin embargo, esta conclusión ha sido criticada, argumentando que las personas obesas reportan que comen más por causas emocionales que los no obesos porque están cumpliendo un rol social (Allison y Heshka, 1993).
2.4. La hipótesis de la restricción
Las investigaciones de campo han demostrado que las personas obesas casi siempre intentan restringir la ingesta de alimentos. Aunque éste es un fenómeno que, en la actualidad, no está limitado a personas obesas, afectando también a otros ámbitos como el estético o el deportivo, donde la restricción calórica, el déficit calórico, o incluso el ayuno, son los términos que se aplican a la población en general que no es obesa, sino que no quiere sentirse como tal o llegar a serlo.
En la hipótesis de la restricción (Herman y Mack, 1975; Herman y Polivy, 1980), se indica que el equilibrio entre el deseo de comer y el esfuerzo por resistir a ese deseo afecta a los comportamientos alimentarios, y la restricción es el esfuerzo cognitivo para resistir ese deseo. Los comedores restringidos se preocupan constantemente por lo que comen y restringen de forma crónica su ingesta de alimentos para evitar engordar. En el otro extremo están los comedores desenfrenados que comen libremente y no se preocupan por su ingesta de alimentos ni por sus consecuencias.
Estos autores también postularon una hipótesis de la desinhibición, según la cual el autocontrol de los comedores restringidos puede liberarse temporalmente mediante eventos perturbadores o desinhibidores que incluyen cogniciones o pensamiento específicos, como comer en exceso; la ingesta de alcohol o los estados emocionales alterados, como la ansiedad o la depresión.
Una revisión de la literatura concluyó que esta hipótesis había sido demostrada (Ruderman, 1986). La mayor parte de la atención se ha centrado en la suposición de que la percepción de haber comido en exceso desinhibe a los comedores restringidos. Esta suposición se ha podido transformar en una variable haciendo que los sujetos comieran con anterioridad antes de una prueba de sabor. En general, los estudios demuestran que hacer una ingesta previa alta en calorías conduce a comer en exceso en los comedores restringidos de forma crónica, ya que tienden a pensar de manera rígida, en forma de todo o nada. También se ha estudiado la influencia del alcohol, pero los resultados no están claros. Se han examinado los efectos de los estados emocionales sobre el consumo de alimentos en comedores restringidos y nos restringidos (Herman y Polivy, 1984), planteando la hipótesis de que las emociones fuertes demandan energía en los comedores restringidos, lo que disminuye temporalmente su motivación para hacer dieta y les permite comer en exceso. Aunque la hipótesis de la restricción predice que cualquier emoción fuerte desinhibe a los comedores restringidos, la investigación se ha centrado principalmente en los efectos de la ansiedad y la depresión en la alimentación (Ruderman, 1986). Aunque se han encontrado resultados contradictorios o difíciles de explicar, este mismo autor pudo llegar a la conclusión de que los estados afectivos negativos, de forma general, aumentan el consumo de los comedores restringidos, pero su impacto en los comedores no restringidos no está tan claro. En un estudio sobre los efectos negativos inducidos por el miedo que se siente al ver una película de terror se produjo el hallazgo de que provocan comer en exceso en sujetos restringidos (Schotte et al., 1990). De nuevo, estos resultados no se encontraron en sujetos no restringidos. En investigaciones de campo también se han podido examinar otros estados emocionales además de la ansiedad y la depresión, alcanzando resultados similares: la exposición a una segmento de una película de comedia desinhibe la ingesta en comedores restringidos (Cools et al., 1992). También se ha podido analizar si la tristeza inducida por una película, caracterizada por una alta valencia negativa y baja aurosal, incrementaba la ingesta en comedores restringidos (Sheppard-Sawyer et al., 2000). Así la exposición a segmentos de películas tristes reducía significamente la ingesta en los comedores no restrictivos, pero la aumentaba de manera no significativa en los comedores restringidos. Ante estos datos, los autores plantearon la hipótesis de que los participantes restringidos pueden no exhibir una alimentación desinhibida cuando se exponen a cambios de humor que no amenazan su autoestima.
3. Emociones y conducta de ingesta
En los seres humanos la conducta alimentaria es compleja y se ve afectada tanto por el estado de ánimo como por las emociones (Lyman, 1982). Como ya se ha señalado en la introducción de este capítulo, el estado de ánimo y las emociones son variables que operan de manera diferente. El estado de ánimo se viene caracterizando por una excitación psicológica en ausencia de estímulo, por lo que conlleva diversos procesos cognitivos, con una duración temporal superior, algunos autores (Macht, 1999) lo han fijado en minutos o más. Por otro lado, las emociones son respuestas psicológicas a corto plazo a estímulos reforzadores, ya sean su naturaleza positiva o negativa.
Los individuos adoptan diferentes comportamientos para regular su estado de ánimo; uno de ellos es el consumo de alimentos. La interacción entre el estado de ánimo, las emociones y la conducta alimentaria es compleja, y supone que las personas regulan sus emociones y su estado de ánimo cambiando tanto las elecciones como las cantidades de alimentos. El estado de ánimo se puede ver afectado por los mecanismos de auto recompensa del consumo de alimentos (Morris y Reilly, 1987). Se prefieren tipos específicos de alimentos bajo algunas condiciones psicológicas debidas a la influencia de los alimentos en la actividad de los centros de recompensa del cerebro. Los alimentos muy sabrosos activan las mismas regiones cerebrales de recompensa y placer que están activas en las adicciones (Volkow et al., 2013). La dopamina, que activa directamente los centros de recompensa y placer, afecta tanto al estado de ánimo como a la ingesta de alimentos (Cantello et al., 1989; Cawley et al., 2013).
De todas las emociones consideradas, un estudio demuestra que las emociones más frecuentes, como la ira o la alegría, tienen una mayor influencia en el apetito y en la elección de alimentos (Macht, 1999). Los hallazgos basados en la medición con cuestionarios del comportamiento humano, estudios de campo y clínicos sugieren un modelo integrador de cinco vías que predice aspectos diferentes de la alimentación emocional. Estos cinco aspectos incluyen: elección de alimentos, consumo de alimentos, pérdida de controles cognitivos, emociones moduladoras de los alimentos y alimentación congruente con las emociones (Macht, 2008). De este modo:
Las emociones inducidas por alimentos, como las que provocan los alimentos ricos en energía, pueden producir ansia acompañadas de respuestas autonómicas (Nederkoorn, Smulders, Jansen, 2000) y también atracones (Jansen, 1998).
Las emociones intensas o el estrés crónico suprimen la ingesta de alimentos (Greeno y Wing, 1994).
Tanto las emociones negativas como las positivas perjudican el control cognitivo de la alimentación y aumentan la ingesta de alimentos. Esto puede deberse a una ruptura de las restricciones autoimpuestas por parte de la persona que hace dieta en presencia de una preocupación más urgente, como el estrés (Herman y Polivy, 1984), o a la demanda de atención creada por el procesamiento de estímulos emocionales que limita la capacidad cognitiva de las personas para mantener su restricción de ingesta de alimentos (Boon, Stroebe, Schut y Jansen, 1998; Ward y Mann, 2000). La mayoría de los estudios sugieren que cuanto más control cognitivo ejerzan las personas sobre su ingesta, más vulnerables serán a comer en exceso.
Las emociones negativas provocan la ingesta para regular las emociones, usando esta conducta como afrontamiento (Buch, 1973; Slochower, 1983). Macht (1999) analiza la hipótesis de la serotonina en la que afirma que las comidas ricas en carbohidratos mejoran el estado de ánimo a través de un aumento de la serotonina cerebral inducida por las comidas (Benton y Donohoe, 1999; Markus et al., 1998) y su efecto endocrino. La hipótesis supone que la ingestión de alimentos ricos en grasas y carbohidratos reduce la actividad suprarrenal del eje hipotálamo-hipofisario y amortigua la respuesta al estrés. Las reacciones afectivas inmediatas a los alimentos sabrosos son el factor más importante en la reducción del impacto del estrés (Macht y Müller, 2007).
Las emociones modulan la conducta de ingesta en congruencia con las características de las emociones. Durante el estado de ánimo negativo, la información verbal negativa se recupera más fácilmente que la información positiva y a la inversa, la información positiva se recupera más fácilmente durante el estado de ánimo positivo. Macht y Müller demostraron que la mejora del estado de ánimo negativo inducido experimentalmente estaba relacionada con la alimentación emocional.
Así dependiendo del estado de las emociones negativas, como la angustia, se desencadena el comer emocional donde la ingesta de alimentos puede aumentar o disminuir dentro de los mismos individuos (Ouwens et al., 2009). El estado emocional también se ha relacionado con la adicción (Parylak et al., 2011).
Los estados de ánimo ansiosos o deprimidos afectan a la elección de alimentos y al metabolismo energético. Comer en exceso, que puede conducir a la obesidad, se asocia con depresión y ansiedad en humanos, lo que también se ha informado en modelos animales (Novick et al., 2005; Sharma y Fulton, 2013). Las personas que experimentan un estado de ánimo deprimido muestran preferencia y consumen “alimentos reconfortantes”, más sabrosos, como un medio para aliviar sus sentimientos negativos (Macht, 2008). Aunque a corto plazo los alimentos sabrosos pueden proporcionar cierto alivio de las emociones negativas y los estados de ánimo, el consumo crónico de alimentos ricos en calorías conduce en última instancia a la obesidad, lo que a su vez promueve vulnerabilidad a la depresión y la ansiedad ( Sharma y Fulton, 2013).
Por otra parte, hay hallazgos que demuestran que la alimentación prolongada con un alto contenido en grasas conduce a estados emocionales negativos, mayor sensibilidad al estrés y niveles alterados de corticosterona basal (Sharma et al., 2012). De esta manera, la emoción negativa afecta a la elección y a la ingesta de alimentos que, a su vez, afecta al estado de ánimo. En modelos de experimentación animal, después de una exposición crónica a una dieta alta en grasas y después de cambiar a dieta normal, los ratones mostraron ansia de sacarosa, alimentos ricos en grasas y un comportamiento similar al de la ansiedad (Sharma et al., 2012). Además, se ha informado de hallazgos similares en el aumento de los signos conductuales y fisiológicos de estados de ánimo ansiosos y deprimidos en humanos, cuando se cambió una dieta con un contenido alto en azúcar y grasa a una dieta normalizada (Avena et al., 2008; Blasio et al., 2013).
3.1. Efectos de las expectativas y la memoria
Aunque está claro que las expectativas juegan un papel importante en la percepción y apreciación de los alimentos y en la elección de los mismos, las investigaciones se han centrado más en aspectos emocionales.
En una investigación en la que se presentaron seis productos para untar de avellana y cacao en tres condiciones (ciego, esperando el paquete e informado) y midieron el gusto y las emociones utilizando el cuestionario EmoSemio que consta de 23 oraciones que describen las emociones. Para dos de sus productos, el agrado esperado y el agrado informado fueron más altos que el agrado bajo la presentación ciega (Spinelli et al., 2015).Estos resultados se utilizan de manera habitual como estrategia de presentación de los productos en restauración y hostelería, propiciando un cambio emocional antes de empezar a comer, aumentando la satisfacción de los clientes.
Por otro lado, las influencias descritas son mucho más débiles que las que se basan en experiencias, en concreto, en situaciones de comidas anteriores y que llevan consigo contenido emocional. Existe una amplia evidencia de que no recordamos con precisión la última comida que hicimos, sino que recordamos inmediatamente la situación anterior en que la comimos, su ambiente o la compañía con la que comimos, o notamos desviaciones de ella como sorpresa o advertencia (Koster et al., 2014; Morin–Audebrand et al., 2012).
4. Estrés y conducta de ingesta
Existe una estrecha interacción entre la comida, el estado de ánimo y el estrés (Benton y Donchoe, 1999; Bast y Berry, 2014). El estrés puede afectar al comportamiento alimentario lo que provoca un aumento o una reducción de la ingesta de alimentos según los tipos de factores estresantes sean externos o psicológicos (Oliver y Wardle, 1999; Yau y Potenza, 2013). De esta forma, el estrés crónico puede conducir a un mayor consumo de alimentos sabrosos y gratificantes que puede derivar en obesidad o en una disminución del apetito que conduce a la pérdida de peso (Cartwright et al., 2003; Tyron et al., 2013). Así, después de la exposición a un estresor, los estudios muestran que la ingesta de alimentos sabrosos reduce los signos de estrés y ansiedad (Pecoraro et at., 2004; Finger et al., 2011 y 2012). La preferencia inducida por el estrés para la ingesta de alimentos sabrosos se observa a menudo en seres humanos (Souquet y Rowland, 1989; Rho et al., 2014). Cabe reseñar que este tipo de comportamiento se extiende también al modelo de experimentación animal (Dallman et al., 2003; Cottone et al., 2009). Esto podría sugerir una vía neurobiológica común para la elección de alimentos durante periodos de estrés, ya sean reales o percibidos.
El nivel de cortisol sérico es una de las medidas asociadas al estado de ánimo. El nivel alterado del mismo, junto con una hiperreactividad del eje hipotalámico-pituitario-suprarrenal (HPA) y la ingesta se asociaría con emociones negativas (Schellenkens et al., 2012).
Se ha observado una superposición de los circuitos neuronales de la ingesta de alimentos y el estrés que probablemente reforzaría el vínculo entre el estrés y el comportamiento alimentario (Maniam y Morris, 2012). En modelos de experimentación con animales, estos circuitos superpuestos del eje HPA que modulan el comportamiento alimentario y el estrés convergen en las neuronas productoras de la hormona corticosterona del núcleo paraventricular. Así, los niveles elevados de glucocorticoides y un eje HPA disfuncional son comunes tanto para el estado de ánimo deprimido como la ingesta excesiva de alimentos.
Como es sabido los glucocorticoides ejercen múltiples efectos sobre las funciones metabólicas, endocrinas, inmunitarias y conductuales. Además, regulan procesos emocionales y de recompensa a través de sus receptores y del circuito límbico. Los glucocorticoides no solo actúan para mantener la homeostasis energética, sino también para modular la actividad del eje HPA y los efectos emocionales y conductuales del estrés (Federoff et al., 2003; Figueiredo et al., 2003).
Bajo estrés fisiológico agudo, el eje HPA se activa con la consiguiente liberación de glucocorticoides, esto conduce a una mayor restauración del equilibrio energético, aumentado la motivación por la comida sabrosa (Piazza y Le Moal, 1997; Dallman, 2010) y acumulando la energía ingerida en exceso en las reservas centrales en el tejido adiposo, lo que conduce a un aumento de peso. De esta manera, el aumento de peso y el estado de ánimo están vinculados a través del eje HPA. En roedores la exposición crónica a la corticosterona conduce a un aumento de la expresión del receptor de glucocorticoides en el cerebro anterior y la amígdala basolateral que da como resultado comportamientos depresivos, similares la ansiedad y un aumento de la agitación (Wei et al., 2004 y Boyle et al., 2005). Por lo tanto, estos hallazgos sugieren que un déficit en la señalización de glucocorticoides en distintas regiones del cerebro puede desempeñar un papel en el trastorno afectivo (Minati, 2014).
Como colofón a lo expuesto, se puede concluir que el estrés afecta significativamente a la ingesta de alimentos tanto en humanos como en animales, lo que promueve alteraciones metabólicas (Block et al., 2009; Manian y Morris, 2012).
5. Ingesta y estado de ánimo
La atención prestada por los investigadores al efecto de los alimentos sobre el estado de ánimo no ha llamado tanto la atención como el efecto del estado de ánimo sobre la elección de alimentos y la ingesta de los mismos. Aparte de la hipótesis de la serotonina mencionada anteriormente, no ha habido otros esfuerzos relevantes para encontrar mecanismos fisiológicos razonables y la hipótesis de la serotonina presenta ciertas limitaciones. En este sentido Haddock y Dill (2000) respaldan la conclusión a la que llega Christensen (1993) “que uno de los pasos bioquímicos requeridos para la ingestión de carbohidratos es que se altere la síntesis de serotonina (Wurtman et al.,1981), lo que sugiere que los efectos de la alteración del estado de ánimo resultantes de la ingestión de carbohidratos probablemente estén mediados por un mecanismo distinto o adicional a la mejora de la síntesis de serotonina o la neurotransmisión”. Además el mecanismo de aumento de la serotonina activado por la ingestión de carbohidratos puede verse fácilmente alterado por la presencia de proteínas en la dieta, y como es sabido las comidas rara vez están libres de proteínas. Finalmente, se ha demostrado que la eficacia de los alimentos ricos en carbohidratos y pobres en proteínas dependía de la propensión al estrés de los sujetos, en concreto, solo se producía en personas con una alta propensión al estrés (Markus et al.,1998 y 2000).
A pesar de que el mecanismo fisiológico subyacente no está bien determinado, el efecto inmediato de los alimentos y especialmente, los carbohidratos y los alimentos dulces, en la reducción de los sentimientos negativos y el estado de ánimo sigue estando bien establecido (Macht y Dettmer, 2006; Macht y Muller, 2007).
Las investigaciones de ensayos en humanos y los modelos de experimentación animal han demostrado que la ingesta de cierto tipo de alimentos respecto a otros influye directamente en los sistemas de neurotransmisores cerebrales, lo que produce efectos sobre el estado de ánimo y el rendimiento al alterar la estructura, la química y la fisiología del cerebro. Como ya se ha comentado en el apartado anterior, el estado de ánimo puede influir en la elección de alimentos y en las expectativas sobre los efectos que estos alimentos pueden producir en función de experiencias previas, fruto del aprendizaje.
Algunos de los alimentos que afectan al estado de ánimo se analizan a continuación:
- El chocolate tiene un fuerte efecto sobre el estado de ánimo, generalmente aumentando los sentimientos agradables y reduciendo la tensión (Osman y Sobal, 2006, Fletcher et al., 2007). Además contiene sustancias químicas como las anandamidas que estimulan el cerebro y dan como resultado un buen estado de ánimo (Ottley, 2000). Aunque los sentimientos negativos también estarían asociados con la ingesta de chocolate en algunas mujeres en régimen de pérdida de peso que experimentan culpa después de comer chocolate (Macht y Dettmer, 2006; Fletcher et al., 2007).
- La cafeína consumida en forma de café o té, no solo tiene efectos estimulantes para mejorar el estado de alerta, la vigilancia y el tiempo de reacción, sino que también aumenta la ansiedad en personas susceptibles (Acquas et al., 2002; Rossi et al., 2010). La cafeína bloquea los receptores de adenosina en el cerebro y puede aliviar dolores de cabeza, la somnolencia y la fatiga. La privación de cafeína a corto plazo, en consumidores habituales produce síntomas de abstinencia (Rogers, 1995).
- Los ácidos grasos omega-3 que se encuentran en varios alimentos, pueden influir en el estado de ánimo, el comportamiento, el neuroticismo y el control de impulsos (Van Straten y Bouvy, 2006; Stahl et al., 2008). Este tipo de ácidos grasos desempeñan un papel en el trastorno depresivo mayor, el trastorno bipolar, la esquizofrenia, el abuso de sustancias y el trastorno por déficit de atención (Young y Martin, 2003; Stahl et al., 2008). Los niveles sanguíneos bajos de ácidos grasos omega-3 poliinsaturados están asociados con la depresión, lo que implica un papel en los trastornos del estado de ánimo (Lombard, 2000; Grosso et al., 2004).
Algunos micronutrientes, como la tiamina, también llamada vitamina B1; el hierro y el ácido fólico, desempeñan un papel en las emociones. Los alimentos que contienen tiamina influyen en los estados de ánimo. Un aumento en el nivel de tiamina aumenta el bienestar, la sociabilidad y los niveles de energía. Las cantidades insuficientes de tiamina se asocian con el deterioro del estado de ánimo y del funcionamiento cognitivo (Benton et al., 1997; Benton y Donohoe, 1999).
La deficiencia de hierro representa uno de los problemas nutricionales más comunes en todo el mundo. La anemia por deficiencia de hierro puede resultar en un estado de ánimo deprimido y problemas de atención (Benton y Donhoe, 1999).
El ácido fólico juega un papel importante en el cerebro. La deficiencia de este ácido se asocia con un estado de ánimo depresivo (Coppen y Bolander Gouaille, 2005; Young, 2007). La evidencia sugiere que un nivel bajo de folato puede ser perjudicial para el estado de ánimo y estar asociado con niveles reducidos del neurotransmisor serotonina (5-hidroxitriptamina; 5-HT) en el líquido cefalorraquídeo (Williams et al., 2005).
En el año 2001 se reconoció por primera vez una ruta potencial de información interna sobre el comportamiento emocional, donde los nutrientes del intestino se transmitían al cerebro mediante el nervio vago que afectaba a las emociones (Zagon, 2001). Sin embargo, la relación de las emociones, la excitación fisiológica y el estado de ánimo en una situación dada depende significativamente del estado motivacional de sujeto (Reid Y Hammersley, 1999) y del rasgo de neuroticismo del individuo que interactúa con el estado de ánimo y la respuesta a los estímulos emocionales (Dess y Edelheit, 1998).
Cierto tipo de dietas pueden desencadenar la patogenia de los trastornos de estado de ánimo y metabólicos. Las dietas como las occidentales que son ricas en grasas saturadas y bajas en ácidos grasos poliinsaturados y monoinsaturados tienden a aumentar la incidencia de estado de ánimo deprimido (Sánchez Villegas et al., 2013).
6. Hormonas implicadas en la ingesta y emociones
Grelina
El eje intestino-cerebro media la comunicación entre estos órganos cuando se trata de apetito, saciedad y homeostasis energética (Cummings y Overdrum, 2007; Suzuki et al., 2010, 2012). Además también se ha informado que las hormonas periféricas regulan el estado de ánimo y la ingesta de alimentos (Tschop et al., 2000; Karra et al., 2013). Es sabido que las señales gastrointestinales como la colecistoquinina (CCK), la bombesina, y el glucagón, entre algunos otros, reducen la ingesta de alimentos, mientras que la excepción es la grelina que aumenta dicha ingesta.
El sistema grelinérgico media los aspectos motivacionales y gratificantes hedónicos no homeostáticos de la ingesta de alimentos a través del circuito dopaminérgico mesolímbico (Dickson et a., 2011; Perelló y Zigman, 2012). Los estudios respaldan la participación en el comportamiento de recompensa alimentaria mediada por estrés (Perelló et al., 2012; Diz-Chaves, 2011). Además numerosos estudios proporcionan un vínculo entre la grelina y los trastornos afectivos, como la depresión y la ansiedad (Shanze et al., 2008; Kluge, 2009). La grelina también alivia la depresión (Kluge et a., 2011).
Serotonina
Además de regular el estado de ánimo y el apetito, la serotonina tiene numerosas funciones ya citadas en los apartados anteriores. Cabe destacar que los niveles de serotonina del intestino y del tracto digestivo constituyen aproximadamente el 80-90% de la serotonina total del cuerpo humano y no siendo el cerebro el órgano principal, a pesar de que serotonina dicta la mayor parte de nuestro estado de ánimo y felicidad (Wurtman y Wurtman, 1989; Benton y Donohoe, 1999).
Las vías centrales de serotonina participan en la regulación del estado de ánimo y modulan los patrones de comidas en términos de calidad y cantidad. La liberación de serotonina de las neuronas serotoninérgicas en el cerebro se rige por la ingesta de alimentos (Shabbir et al. 2013). El aminoácido esencial triptófano que proviene de los alimentos es el precursor de la síntesis de serotonina (Prasad, 1998). La ingestión de carbohidratos aumenta la proporción plasmática de triptófano a otros aminoácidos neutros grandes, lo que conduce a un aumento de la síntesis de serotonina en el cerebro y a aliviar el estado de ánimo deprimido. Tal es el caso del ansia por carbohidratos durante la depresión que a menudo conduce a la obesidad y viceversa (Pepino et al., 2009; Shabbir et al., 2013). Esto se observa durante el estrés, la depresión invernal o en personas que intentan dejar de fumar. La nicotina aumenta la secreción de serotonina en el cerebro y su abstinencia conduce a la depresión (Wallin y Rissanen, 1994; Wurtman y Wurtman, 1996). La serotonina cerebral desempeña un papel en la fisiología de la depresión ya que los tratamientos con fármacos potenciadores de la serotonina alivian la depresión en el trastorno afectivo estacional (Wurtman, 1993). Estos hallazgos han sugerido que la ingesta de carbohidratos por parte de pacientes con síndrome premenstrual y el trastorno afectivo estacional alivia los síntomas depresivos a través de un aumento de la actividad serotoninérgica central (Cizza et al., 2005; Miller, 2005).
Por otro lado, la investigación ha demostrado que las personas que hacen dieta restrictivas tienden a tener un ánimo deprimido a medida que los niveles de serotonina se reducen debido a la disminución de la ingesta de carbohidratos (Huether et al., 1997)
Leptina
Diversas investigaciones han puesto de manifiesto que los niveles bajos de leptina están asociados con el estado de ánimo deprimido en humanos y en comportamientos similares a la depresión en roedores. (Kraus et at., 2001; Lawson et al., 2012). El efecto antidepresivo de la leptina en situaciones de insuficiencia de nivel de leptina o con resistencia a la leptina sugiere que la hormona contribuye al estado de ánimo alterado (Lu, 2007). El aumento de grasa visceral y la dislipidemia están asociados con varios cambios endocrinos y metabólicos que se relacionan con el control del SNC de los estados emocionales y el estado de ánimo (Hryhorczuk et al., 2013). Como glándula endocrina, el tejido adiposo secreta numerosas hormonas peptídicas que actúan sobre el cerebro y los tejidos periféricos para regular el metabolismo y el comportamiento. La leptina circula, como es sabido, en proporción a la masa grasa (Maffei et al., 1995). La hormona se ha relacionado, entre otros, con la depresión humana y se ha demostrado que en roedores tiene efectos antidepresivos y ansiolíticos (Asakawa et al., 2003; Lawson et al., 2012). Hay que destacar que en relación a los niveles en plasma de la hormona existen hallazgos contradictorios en humanos. Es de esperar que futuras investigaciones determinen la relación entre la leptina y el estado de ánimo.
Sobre esta cuestión, los estudios con roedores han proporcionado los hallazgos más concluyentes. La leptina modula el eje HPA y los ratones que carecen de leptina muestran un comportamiento similar a la depresión (Collin et al.,2000; Guo et al., 2012). Por el contrario, el estrés crónico leve e impredecible en ratas activa el eje HPA y conduce a conductas de tipo depresivo que se relaciona con la disminución del nivel de leptina sérica (Ge et al., 2013). La activación de los receptores de leptina (LepRb) en los loci del mesencéfalo y el prosencéfalo afectan a los procesos emocionales. La eliminación genética de LepRb en el hipocampo da como resultado un fenotipo similar a la depresión, que se reduce con la administración de leptina en el hipocampo, mostrando así efectos antidepresivos (Asakawa et al., 2003; Guo et al., 2013). La pérdida de LepRb específicamente de las neuronas glutamatérgicas del cerebro anterior provoca un comportamiento depresivo que no afecta a la ansiedad (Guo et al., 2012). La liberación de dopamina inducida por estrés también se asocia con niveles altos de leptina (Burghardt et al., 2012). La eliminación selectiva de LepRb de las neuronas de dopamina del mesencéfalo da como resultado un aumento del comportamiento a la ansiedad, pero no al comportamiento depresivo (Liu et al., 2011). La señalización de LepRb en los núcleos límbico y prefrontal media acción antidepresiva de la leptina. Sin embargo, la leptina en las neuronas dopaminérgicas del mesencéfalo ventral y en el núcleo central de las amígdala ejerce la la señalización de las acciones ansiolíticas de la leptina, actuando así en diferentes regiones del cerebro y con distintos efectos.
Adiponectina
Los niveles bajos de otra hormona derivada del tejido adiposo, la adiponectina, se han relacionado con homeostasis energética, los trastornos metabólicos y la depresión, entre otros. Aunque hay informes contradictorios de asociaciones positivas y negativas de los niveles de adiponectina con trastornos del estado de ánimo y los trastornos depresivos.
Los ratones expuestos a la derrota social, definida como una lucha fallida en función de las expectativas, muestran niveles bajos de adiponectina, comportamientos depresivos inducidos por estrés y el eje HPA deteriorado. Por el contrario, la administración central de adiponectina tiene efectos antidepresivos (Liu et al, 2012). Así se observa en ratones un vínculo entre los niveles de adiponectina en plasma y la depresión, si bien en humanos los resultados se muestran más ambiguos.
7. Epigenética
La interacción de los genes y el medio ambiente se ha asociado con los trastornos del estado de ánimo. La exposición a alimentos muy sabrosos, ricos en grasas y carbohidratos, induce al antojo, definido como un impulso vehemente. En ambientes determinados, la exposición repetitiva a opciones de alimentos altamente apetecibles aumenta la probabilidad de adicción a ese tipo de alimentos y a comer en exceso (Singh, 2014).
La variabilidad individual en numerosas regiones codificantes y no codificantes de proteínas en el genoma podría estar relacionada con trastornos alimentarios y trastornos afectivos. Los mecanismos epigenéticos de la metilación del ADN, la edición de ARN, la modificación postraduccional de las histonas y los ARN no codificantes regulan la expresión génica sin cambiar la secuencia del ADN en respuesta a cambios en las variables ambientales internas y externas. La epigenética en el contexto alimentario es interesante, ya que permitiría responder a numerosas preguntas, incluidos los factores de riesgo potenciales, como la nutrición materna y el estrés que alteran el riesgo de trastornos en la descendencia.
A modo de ejemplo de esta multiplicidad de variables asociadas, se destaca el receptor 5HT2CR de la serotonina que se ha asociado con trastornos afectivos, el estrés y otros síndromes. La edición de ARN del 5HT2CR altera muchas facetas de la señalización de la serotonina a través de 24 isoformas de receptores diferentes. Estas isoformas editadas están en distinta proporción en diferentes regiones del cerebro lo que sugiere un papel importante en la vinculación con el estado de ánimo, y la ingesta de alimentos (Singh, 2014).
8. La ingesta emocional
El término emotional eating se viene utilizando en la literatura científica desde los años 80 del siglo pasado en el contexto de los estudios sobre la obesidad (Lowe, 1983) que ya han sido tratados en apartados anteriores. En estos estudios se reporta una fuerte relación entre la ingesta y las emociones negativas o los eventos estresantes en la vida. La ingesta emocional ocurre de forma episódica y no de forma regular; se hace en secreto, se asocia con diferentes emociones y se caracteriza por el uso de alimentos ricos en calorías y carbohidratos (Ganley, 1989). Se ha descubierto que la ingesta emocional es más frecuente cuando las personas están solas, cuando la comida es una cena o un refrigerio y cuando la comida se come en casa en comparación a cuando se realiza fuera de la misma (Baumeister et al., 1994). La alimentación emocional prevalece en las distintas clases sociales y sexos. Los estudios informan consistentemente que la ingesta emocional es, más a menudo, precipitada por emociones negativas como la ira, la depresión, el aburrimiento, la ansiedad y la soledad y a menudo, tiene una relación episódica con periodos estresantes de la vida. Una revisión de campo concluyó que en sujetos con obesidad mórbida que buscan tratamiento, la ingesta emocional parece ser muy común (Ganley, 1989).
La ingesta emocional no es un trastorno alimentario aislado, sino un comportamiento alimentario que está influenciado por otros comportamientos, como el estrés, las emociones y los sentimientos individuales en relación con la alimentación. Sin embargo la ingesta emocional, a diferencia de los trastornos alimentarios específicos, no está relacionada con una falta total de control de la cantidad y la calidad de los alimentos consumidos (Grajek et al., 2022). La alimentación emocional o de comodidad, así como la alimentación inducida por el estrés, conduce a la predisposición a comer en respuesta a emociones negativas, siendo los alimentos preferidos principalmente ricos en energía, pobres en nutrientes y sabrosos (Godet et al., 2022; Pla-Sanjuanelo et al., 2015; Lu et al., 2016). Estos tipos de alimentación actúan como un mecanismo de afrontamiento para controlar y disminuir las emociones negativas, como el estado de ánimo deprimido, la ansiedad y el estrés (Sambal et al., 2021). En cuanto a la prevalencia del comer emocional, con base en la encuesta Stress in America realizada por la Asociación Americana de Psicología, el 38% de los adultos afirmaron haber estado implicados en el comer emocional durante el último mes, con un 49% de ellos participando semanalmente (Rahme et al., 2021). Además, con frecuencia se considera un factor principal en modelos de trastornos alimentarios y actitudes patológicas hacia la alimentación, como comer en exceso y atracones (Dakanalis et al., 2017; Wolz et al., 2021), y podría provocar trastornos psicológicos y problemas de salud importantes (Pentikäinen et al., 2018). Por lo tanto, es importante interpretar sus mecanismos fundamentales (Sambal et al., 2021).
La alimentación emocional tiene una relación positiva con el aumento de la ganancia de peso a lo largo del tiempo y la dificultad para perder peso. Esto se puede atribuir al hecho de que los comedores emocionales son más propensos a un mayor consumo de alimentos azucarados y ricos en grasas, comen en respuesta a factores estresantes y comen bocadillos con más frecuencia en comparación con los comedores no emocionales (Braden et al., 2018) . En particular, un análisis transversal de la cohorte NutriNet-Santé realizado en 7.378 hombres y 22.862 mujeres informó que una mayor alimentación emocional se relaciona con un consumo elevado de refrigerios ricos en energía, como alimentos dulces y grasos, y estas asociaciones fueron predominantemente más fuertes en mujeres que sufren de depresión (Camilleri et al., 2014). Además, se ha informado de una asociación entre el consumo excesivo de alcohol y el comer emocional (Azevedo et al., 2021). Estos hábitos alimenticios combinados con un mayor peso corporal ponen a los comedores emocionales en un mayor riesgo de diabetes y enfermedades del corazón (Frayn et al., 2018). Además, los comedores emocionales activan la relación entre el motivo de comer y la recompensa, lo que respalda la evidencia de que la comida ejerce un efecto de recompensa y, por lo tanto, puede aliviar o disminuir las condiciones negativas del estado de ánimo (Godet et al., 2022). Además, los comedores emocionales con frecuencia sienten emociones negativas con respecto a su apariencia física justo después de comer. Prácticamente todas las personas han experimentado el impacto de las emociones en las actitudes alimentarias. Por el contrario, el comportamiento alimentario también puede afectar la imagen corporal, lo que resulta en insatisfacción corporal, lo que implica una autoevaluación negativa relacionada con el cuerpo de una persona (Rahme et al., 2022).
Las posibles causas de la ingesta emocional son una restricción dietética alta, una conciencia interoceptiva deficiente, alexitimia, desregulación de las emociones y un eje de estrés hipotalámico pituitario suprarrenal (HPA) invertido. La alimentación emocional puede ser el resultado de una crianza inadecuada o sentimientos depresivos en interacción con la susceptibilidad genética. También hay pruebas sólidas de que la ingesta emocional es un mediador entre la depresión y la obesidad (van Strien, 2018).
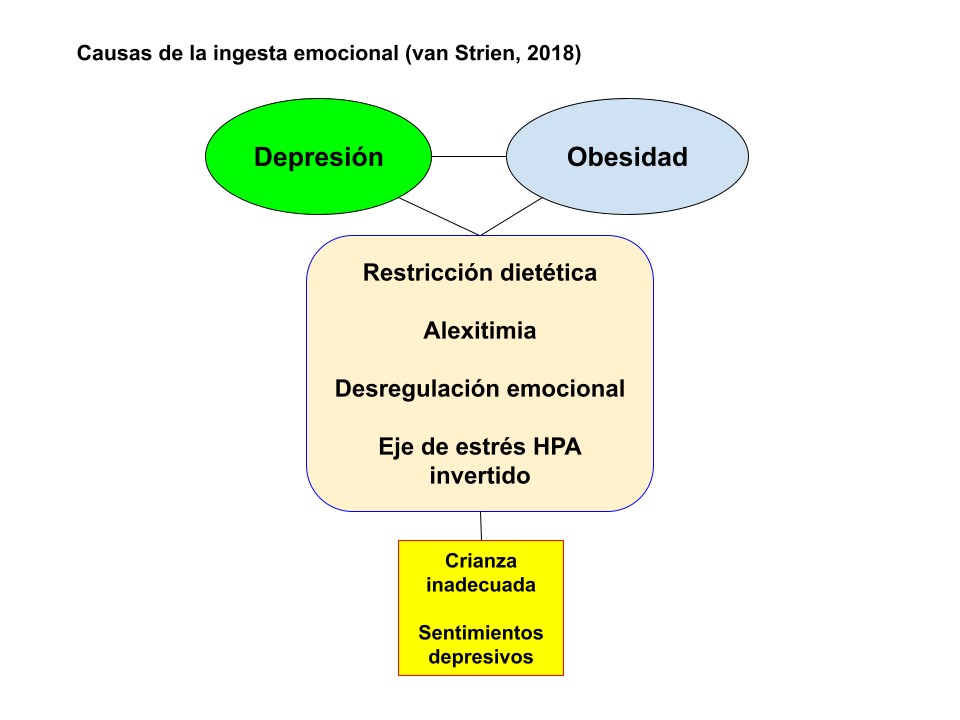
Por otro lado, el estrés y las condiciones emocionales afectan al comportamiento alimentario (Dakanalis et al., 2014). El estrés y el estado de ánimo negativo pueden influir inversamente en el apetito, lo que lleva a varias personas a comer más y a otras a comer menos (Evers et al., 2018; Dakanalis et al., 2013). Estos comportamientos, conocidos como sobrealimentación emocional y desnutrición emocional, se han asociado con el inicio de complicaciones relacionadas con el peso corporal y trastornos alimentarios (Dakanalis et al., 2016 y 2017). Sin embargo, en un subgrupo significativo de personas de ambos sexos, las emociones negativas y el estrés los hacen comer en exceso (Buja et al., 2022; Konttinent et al., 2010). Así el funcionamiento emocional anormal significa dificultad para reconocer, comprender y afrontar adecuadamente las emociones y una experiencia a largo plazo de niveles elevados de emociones dañinas (Czepczor-Bernat, et al., 2020). En particular, la afectividad negativa elevada, la insatisfacción corporal, la auto-objetivación y la baja autoestima fueron predictivos de participación persistente en atracones regulares y conductas compensatorias de peso inapropiadas. La auto-objetivación, considerada como pensar y monitorear la apariencia externa del cuerpo desde una perspectiva de tercera persona, surgió como el mayor contribuyente tanto del inicio como de la persistencia de todos los síntomas conductuales (Dakanalis et al., 2016 y 2017).
En relación a las personas que hacen dieta, el estrés y las emociones negativas pueden desinhibir o liberar el autocontrol de las personas con respecto a la restricción autoimpuesta de la ingesta de alimentos. Cuando se come de menos, el cuerpo no puede distinguir la restricción alimentaria autoimpuesta de la escasez real de alimentos y actúa como si estuviera en estado de inanición: la tasa metabólica se ralentiza (anabolismo) y el hambre y el apetito aumentan (Goldsmith et at., 2010). El resultado es que las personas que hacen dieta desarrollan intensos sentimientos de privación, porque en realidad la sensación de deprivación es doble, por una lado está la lucha interior se manifiesta contra las sensaciones de su propio metabolismo: falta de energía, cansancio, dificultades cognitivas; y por otro, la sensación de hambre que puede llegar a ser incluso dolorosa. De hecho, cuando estaban bajo estrés o después de la inducción de emociones negativas, las personas que hacían dieta mostraron en varios experimentos una mayor ingesta de alimentos en comparación con las personas que no hacen dieta (Heatherton et al., 1991; Herman y Polivy,1975).
La alimentación emocional también puede ocurrir independientemente de la dieta. Puede ser el resultado de una conciencia interoceptiva deficiente, una confusión de estados internos de hambre y saciedad con síntomas fisiológicos asociados con las emociones (Bruch, 1973), alexitimia o estrategias de regulación emocional deficientes. Alternativamente, comer emocionalmente también se ha asociado con una respuesta de estrés invertida del eje HPA (Van Strien, 2013; Dalman, 2010).
Se ha sugerido que la respuesta antinatural de comer emocionalmente se adquiere como posible resultado de las prácticas de crianza que socavan el desarrollo psicológico y emocional del niño (Snoek, 2007). Cuando la respuesta del cuidador es continuamente inapropiada, ya sea negligente, excesivamente protectora o incluso manipuladora u hostil, el resultado puede ser que el niño desarrolle una conciencia interoceptiva deficiente (conciencia deficiente de los sentimientos de hambre y saciedad), altos grados de alexitimia, definida como la dificultad para identificar sentimientos y describirlos a otras personas (Taylor, 1996), y dificultades para regular adecuadamente sus emociones (Jaffe, 2009; Vandewalle, 2016).
De hecho, la conciencia interoceptiva deficiente y la alexitimia alta se asociaron positivamente con la alimentación emocional autoinformada (McDowell et al., 2002; Van Strien et al., 2005). La alexitimia también moderó la asociación entre el estrés y la ingesta de alimentos, y las mujeres con un alto grado de alexitimia mostraron la respuesta atípica al estrés de comer lo mismo o incluso un poco más después del estrés, medido como variable en la creencia de tener que dar un discurso frente a una audiencia evaluadora (Van Strien y Ouwens, 2007). También se demostró que las estrategias de regulación emocional deficientes, como la supresión de las emociones o las estrategias de afrontamiento desadaptativas, como la confianza en el afrontamiento orientado a las emociones y la evitación del estrés por distracción, se asocian positivamente con la alimentación emocional (Spoor et al., 2007; Lu et al., 2016).
Un posible mecanismo subyacente a la alimentación emocional puede ser una respuesta de estrés invertida del eje HPA como posible resultado del estrés crónico temprano en la vida. En lugar de responder al estrés con hiperactivación y los síntomas neurovegetativos típicos, como la pérdida de apetito, el eje HPA puede responder al estrés con una hipoactividad del eje HPA y los síntomas neurovegetativos (atípicos) de aumento del apetito y aumento de peso (Gold et a., 2012). En esta línea de pensamiento, los comedores emocionales no perciben la típica reducción del hambre posterior al estrés, sino que tienen una sensación de hambre similar o mayor después del estrés se llevó a cabo en pacientes con trastorno por atracón compulsivo, una afección que está estrechamente relacionada con un alto nivel de alimentación emocional (Gluck et al., 2004). Un eje HPA reducido que funcione en respuesta al estrés también explicaría por qué los comedores emocionales son más receptivos al valor de refuerzo de los alimentos y usan los alimentos como estrategia para mitigar de forma temporal, los efectos del estrés y las emociones negativas (Haedt-Matt et al., 2014).
En algunos estudios, se demostró que las experiencias adversas de crianza temprana en la vida, particularmente cuando involucran relaciones entre padres e hijos, tienen efectos duraderos en los sistemas neurológicos que responden al estrés (Yehuda et al., 2000; Cicchetti y Rogosch, 2001). Además, también se ha observado que los comedores emocionales responden al estrés agudo con un aumento en el cortisol (Epel et al., 2007; Rapopow et al., 2010), y en otro estudio se encontró que las mujeres que experimentaron estrés crónico, porque eran cuidadoras de niños con enfermedades crónicas, tenían puntajes más altos en una escala de alimentación emocional y mostraron una actividad del eje HPA amortiguada en comparación con mujeres sin tal estrés crónico (Tomiyama et al., 2011).
La hipótesis de que los comedores emocionales, con una actividad de estrés del eje HPA reducida, respuesta de estrés de cortisol embotada, mostrarían la mayor ingesta de alimentos después del estrés fue confirmada (van Strien et al., 2013). En la prueba se utilizó como estresor el TSST (Trier Social Stress Test) que induce el estrés al exigir a los participantes que hagan una presentación estilo entrevista, seguida de una prueba de aritmética mental sorpresa, frente a un grupo de entrevistadores que no proporciona retroalimentación ni estímulo (Kirschbaum et al., 1993). También en otros dos estudios hubo evidencia de que una respuesta de estrés del eje HPA contundente condujo a una mayor ingesta de alimentos (Tyron et al.,2013; Wingenfeld et al., 2017). Sin embargo, aún no se ha resuelto si una respuesta de estrés de cortisol atenuada es realmente una causa o más bien una respuesta a la alimentación emocional (van Strien et al., 2013). También se ha concebido que una respuesta de estrés de cortisol atenuada es secundaria a la alimentación emocional, como resultado de la regulación adaptativa a la baja del HPA, en respuesta a la sobrealimentación frecuente (Dalman, 2010).
8.1. La ingesta emocional durante la adolescencia
La alimentación emocional es muy frecuente en mujeres adultas con sobrepeso u obesidad (van Strien et al., 2009 y 2012). En los niños por el contrario, la prevalencia de la alimentación emocional es muy baja (Sleddens et al., 2008; van Strien y Oosterveld, 2009). Esto sugiere que la mayoría de los niños pequeños muestran la respuesta natural de perder el apetito en respuesta a las emociones negativas o al estrés y que la alimentación emocional surge en la transición entre la niñez y la edad adulta: el período de la adolescencia, posiblemente como resultado de la activación de los estrógenos en la pubertad (Kumb, 2013; Klumb et al., 2016). Dos estudios prospectivos en familias holandesas con dos hijos adolescentes abordaron la aparición de la alimentación emocional en la adolescencia (van Strien et al., 2010).
En el primer estudio (n = 279), se evaluó la aparición del comer emocional en relación con la interacción entre la práctica parental inadecuada de control psicológico (p. ej., 'Mi padre (madre) me hace sentir culpable cuando fracaso en la escuela) y el efecto dopaminérgico (sensibilidad a la recompensa) en un polimorfismo en el gen del receptor de dopamina D2 (DRD2 ) (van Strien eta al., 2010). Llevar el alelo A1 del polimorfismo Taq1A del gen DRD2 (rs1800497) se asocia con una disponibilidad reducida del receptor DRD2 en el cerebro (Gluskin y Mickey, 2016). En el receptor DRD2 se encontró que el genotipo modera la asociación entre el control psicológico materno y paterno y el posible aumento de la alimentación emocional. Las niñas y los niños adolescentes mostraron un aumento en el comer emocional después de 4 años en relación con un alto control psicológico parental al inicio solo si portaban al menos un alelo DRD2 A1.
En el segundo estudio, se evaluó la aparición de la alimentación emocional en relación con la interacción entre los sentimientos depresivos y un sistema cerebral serotoninérgico, a saber, los polimorfismos en el gen del transportador de serotonina SLC6A4/5-HTT (van Strien et al., 2010). El alelo corto del polimorfismo 5-HTTLPR en el gen transportador de serotonina se asocia con una menor actividad de la serotonina (Harari y Holmes, 2005). Los niveles más bajos de actividad de la serotonina se asocian con un aumento del apetito y del peso corporal (Leibovitz SF, Alexander, 1998). De hecho, el gen transportador de serotonina moderó la asociación entre los sentimientos depresivos y el aumento de la alimentación emocional después de 4 años. En los hermanos menores (que tenían 13 años al inicio del estudio) (n =286), esto fue cierto tanto para las niñas como para los niños. En el hermano mayor (que tenía 15 años al inicio del estudio) (n =298), esto solo fue cierto para las niñas, posiblemente porque las niñas de esta edad tienen una mayor prevalencia tanto de depresión como de alimentación emocional (van Strien et al., 2010), o debido a las diferencias de sexo en las interacciones con el polimorfismo 5-HTTLPR (Gressier et al., 2016).
En ambos estudios, no hubo efectos principales de los polimorfismos. Solo en interacción con la crianza negativa o los sentimientos depresivos, predijeron aumentos futuros en la alimentación emocional. Al observar más de cerca los efectos de interacción, respaldan la afirmación del marco de 'genes de placidez': la noción de que los 'genes de riesgo' están asociados con una mayor sensibilidad al medio ambiente (Belsky et al., 2009). Los adolescentes con al menos un gen de riesgo (al menos un DRD2 alelo A1) que crecieron bajo la experiencia de crianza favorable de no experimentar el control psicológico de los padres mostraron el aumento más bajo de alimentación emocional después de 4 años y parecían ir incluso mejor que los adolescentes que portaban el genotipo A2/A2. Lo mismo sucedió con las adolescentes jóvenes (13 años) y las adolescentes mayores (15 años) que combinaban portar al menos un alelo corto 5-HTTLPR con sentimientos depresivos bajos. La noción de genes de flexibilidad también abre perspectivas para la prevención, es decir, que a las personas con 'genes de riesgo' les puede ir mejor cuando el entorno es favorable. Una conclusión similar y la relevancia de la hipótesis de la "susceptibilidad diferencial" para la obesidad se puede encontrar en una revisión posterior (Molle et al., 2017). El resultado de esta revisión sugiere que la hipótesis de susceptibilidad diferencial de Belsky también se aplica a la interacción GxE en los estilos de alimentación y la obesidad.
REFERENCIAS
Acquas, E., Tanda, G., and Di Chiara, G. (2002). Differential effects of caffeine on dopamine and acetylcholine transmission brain areas drug-naive and caffeine-pretreated rats. Neuropsychopharmacology 27, 182–193.
Allison, D. B., Heshka, S., (1993). Emotion and eating in obesity? A critical analysis. International Journal of Eating Disorders 13, 289/295.
Asakawa, A., Inui, A., Inui, T., Katsuura, G., Fujino, M. A., and Kasuga, M. (2003). Leptin treatment ameliorates anxiety in ob/ob obese mice. J. Diabetes Complicat. 17, 105–107.
Avena,N. M., Rada, P., and Hoebel, B. G. (2008). Evidence For Sugar Addiction: behavioral and neurochemical effects of intermittent, excessive sugar intake. Neurosci.Biobehav. Rev. 32, 20–39.
Azevedo, L. D. S., de Souza, A. P. L., Ferreira, I. M. S., Lima, D. W. D. C., & Pessa, R. P. (2021). Binge eating and alcohol consumption: an integrative review. Eating and weight disorders : EWD, 26(3), 759–769.
Bast, E.S., and Berry, E. M. (2014). Laugh away the fat? Therapeutic humor in the control of stress-induced emotional eating. Rambam Maimonides Med.J 5,e0007.
Baumeister, R. F., Heatherton, T. F., Tice, D. M., (1994). Losing Control: How and How People Fail at Self-Regulation. Academic Press, Inc, San Diego, CA.
Belsky, J., Jonassaint, C., Pluess, M., Stanton, M., Brummett, B., & Williams, R. (2009). Vulnerability genes or plasticity genes?. Molecular psychiatry, 14(8), 746–754.
Benton, E., & Donohoe, R. T. (1999). The effects of nutrients on mood. Public Health Nutrition, 2, 403–409.
Benton, D., Griffiths, R., and Haller, J. (1997). Thiamine Supplementation mood and cognitive functioning. Psychopharmacology (Berl.) 129, 66–71.
Blasio,A., Steardo, L., Sabino, V., and Cottone, P. (2013). Opioid system the medial prefrontal cortex mediates binge-like eating. Addict.Biol. 19,652–662.
Block, J. P. ,He, Y., Zaslavsky, A. M., Ding, L., and Ayanian, J. Z. (2009). Psychosocial stress and change in weight among US adults. Am. J. Epidemiol. 170, 181–192.
Boon, B., Stroebe,W., Schut, H., & Jansen, A. (1998). Food for thought: Cognitive regulation of food intake. British Journal of Health Psychology, 3, 27–40.
Boyle, M. P. ,Brewer, J. A., Funatsu, M., Wozniak, D. F., Tsien, J. Z., Izumi, Y., et al. (2005). Acquired deficit of forebrain glucocorticoid receptor produces depression-like changes in adrenal axis regulation and behavior. Proc. Natl. Acad. Sci. U.S.A. 102, 473–478.
Braden, A., Musher-Eizenman, D., Watford, T., & Emley, E. (2018). Eating when depressed, anxious, bored, or happy: Are emotional eating types associated with unique psychological and physical health correlates?. Appetite, 125, 410–417.
Bruch, H. (1973). Eating Disorders: Obesity, Anorexia Nervosa and the Person Within. Basic Books, New York.
Buja, A., Manfredi, M., Zampieri, C., Minnicelli, A., Bolda, R., Brocadello, F., Gatti, M., Baldovin, T., & Baldo, V. (2022). Is emotional eating associated with behavioral traits and Mediterranean diet in children? A cross-sectional study. BMC public health, 22(1), 1794.
Burghardt, P. R., Love, T. M., Stohler, C. S., Hodgkinson, C., Shen, P. H., Enoch, M. A., et al. (2012). Leptin regulates dopamine responses to sustained stress in humans. J. Neurosci. 32, 15369–15376.
Camilleri, G. M., Méjean, C., Kesse-Guyot, E., Andreeva, V. A., Bellisle, F., Hercberg, S., & Péneau, S. (2014). The associations between emotional eating and consumption of energy-dense snack foods are modified by sex and depressive symptomatology. The Journal of nutrition, 144(8), 1264–1273.
Canetti, L., Bachar, E., and Berry, E. M. (2002). Food And Emotion. Behav. Processes. 60, 157–164.
Cantello, R., Aguggia, M., Gilli, M., Delsedime, M., Chiardo Cutin, I., Riccio, A., et al. (1989). Major depression in Parkinson disease and mood response to intravenous methylphenidate: possible role of the“hedonic”dopamine synapse. J. Neurol.Neurosurg.Psychiatry. 52, 724–731.
Cartwright, M., Wardle, J., Steggles, N., Simon, A. E., Croker, H., and Jarvis, M. J. (2003). Stress and dietary practices in adolescents. Health Psychol. 22, 362–369.
Cawley, E. I., Park, S., Aan Het Rot, M., Sancton, K., Benkelfat, C., Young, S. N., et al. (2013). Dopamine And Light: dissecting effects on mood and motivational states in women with subsyndromal seasonal affective disorder. J. Psychiatry Neurosci. 38, 388–397.
Christensen, L. (1993). Effects of eating behaviour on mood: A review of the literature. International Journal of Eating Disorders, 14(2), 171–183.
Cicchetti, D., & Rogosch, F. A. (2001). Diverse patterns of neuroendocrine activity in maltreated children. Development and Psychopathology, 13(3), 677–693.
Cizza, G., Skarulis, M., & Mignot, E. (2005). A Link Between Short Sleep and Obesity: Building the Evidence for Causation. Sleep, 28, 1217-1220.
Cools, J., Schotte, D.E., & McNally, R.J., (1992). Emotional arousal and overeating in restrained eaters. Journal of Abnormal Psychology 101, 348/351.
Coppen, A., & Bolander-Gouaille, C. (2005).Treatment of depression: time to consider folic acid and vitamin B12. J. Psychopharmacol. 19, 59–65.
Collin, M., Hakansson-Ovesjö, M. L., Misane, I., Ogren, S.O., & Meister, B. (2000). Decreased 5-HT transporter mRNA in neurons of the dorsal raphe nucleus and behavioral depression in obese leptin-deficient ob/ob mouse. Brain Res. Mol. Brain Res. 81, 51–61.
Cottone, P., Sabino, V., Steardo, L., & Zorrilla, E. P. (2009). Consummatory, anxiety-related and metabolic adaptations in female rats with alternating access to preferred food. Psychoneuroendocrinology 34, 38–49.
Cummings, D. E., & Overduin, J. (2007). Gastrointestinal regulation of food intake. J. Clin. Invest. 117, 13–23.
Czepczor-Bernat, K., Brytek-Matera, A., Gramaglia, C., & Zeppegno, P. (2020). The moderating effects of mindful eating on the relationship between emotional functioning and eating styles in overweight and obese women. Eating and weight disorders : EWD, 25(4), 841–849.
Dakanalis, A., Clerici, M., Caslini, M., Gaudio, S., Serino, S., Riva, G., & Carrà, G. (2016). Predictors of initiation and persistence of recurrent binge eating and inappropriate weight compensatory behaviors in college men. The International journal of eating disorders, 49(6), 581–590.
Dakanalis, A., Clerici, M., Bartoli, F., Caslini, M., Crocamo, C., Riva, G., & Carrà, G. (2017). Risk and maintenance factors for young women's DSM-5 eating disorders. Archives of women's mental health, 20(6), 721–731.
Dakanalis, A., Timko, C. A., Carrà, G., Clerici, M., Zanetti, M. A., Riva, G., & Caccialanza, R. (2014). Testing the original and the extended dual-pathway model of lack of control over eating in adolescent girls. A two-year longitudinal study. Appetite, 82, 180–193.
Dakanalis, A., Zanetti, M. A., Clerici, M., Madeddu, F., Riva, G., & Caccialanza, R. (2013). Italian version of the Dutch Eating Behavior Questionnaire. Psychometric properties and measurement invariance across sex, BMI-status and age. Appetite, 71, 187–195.
Dalle Molle, R., Fatemi, H., Dagher, A., Levitan, R. D., Silveira, P. P., & Dubé, L. (2017). Gene and environment interaction: Is the differential susceptibility hypothesis relevant for obesity? Neuroscience and biobehavioral reviews, 73, 326–339.
Dallman, M. F. (2010). Stress-induced obesity and the emotional nervous system. Trends Endocrinol. Metab. 21, 159–165.
Dallman, M. F., Pecoraro, N., Akana, S. F. ,La Fleur, S. E., Gomez, F. ,Houshyar, H., et al. (2003). Chronic stress and obesity: a new view of“comfort food. Proc. Natl. Acad. Sci. U.S.A. 100, 11696–11701.
Damasio, H., Grabowski, T., Frank, R., Galaburda, A. M., & Damasio A. R. (1994). The return of Phineas Gage: clues about the brain from the skull of a famous patient. Science. May 20; 264(5162):1102-5.
Davis, R., Freeman, R. J., & Solyom, L., (1985). Mood and food: an analysis of bulimic episodes. Journal of Psychiatric Research 19, 331/335.
Dess, N. K., & Edelheit, D. (1998). The bitter with the sweet: the taste/stress/temperament nexus. Biol. Psychol. 48, 103–119.
Dickson, S. L., Egecioglu, E., Landgren, S., Skibicka, K. P. ,Engel, J. A., & Jerlhag, E. (2011). The role of the central ghrelin system in reward from food and chemical drugs. Mol. Cell. Endocrinol. 340, 80–87.
Diz-Chaves, Y. (2011). Ghrelin,appetite regulation,and food reward: interaction with chronic stress. Int. J. Pept. 2011:898450.
Ekman, P. (1972). Universals and cultural differences in facial expressions of emotions. Darwin and facial expression: A century of research in review. Ekman, P. (ed.), New York, 169-222.
Epel, E., Jimenez, S., Brownell, K., Stroud, L., Stoney, C., & Niaura, R. (2004). Are stress eaters at risk for the metabolic syndrome?. Annals of the New York Academy of Sciences, 1032, 208–210.
Evers, C., Dingemans, A., Junghans, A. F., & Boevé, A. (2018). Feeling bad or feeling good, does emotion affect your consumption of food? A meta-analysis of the experimental evidence. Neuroscience and Biobehavioral Reviews, 92, 195–208.
Fedoroff, I., Polivy, J., & Herman, C. P. (2003). The specificity of restrained versus unrestrained eaters responses to food cues: general desire to eat, or craving for the cued food? Appetite 41, 7–13.
Figueiredo, H. F., Bodie, B. L., Tauchi, M., Dolgas, C. M., & Herman, J. P. (2003). Stress integration after acute and chronic predator stress: differential activation of central stress circuitry sensitization of the hypothalamo-pituitary-adrenocortical axis. Endocrinology 144, 5249–5258.
Finger, B. C., Dinan, T. G., & Cryan, J. F. (2011). High-fat diet selectively protects against the effects of chronic socialstressinthemouse. Neuroscience 192, 351–360.
Finger, B. C., Dinan, T. G., & Cryan, J. F. (2012). The temporal impact of chronic intermittent psychosocial stress on high-fat diet-induced alterations bodyweight. Psychoneuroendocrinology 37, 729–741.
Fletcher, B. C., Pine, K. J., Woodbridge, Z., & Nash, A. (2007). How visual images of chocolate affect the craving and guilt of female dieters. Appetite 48, 211–217.
Frayn, M., Livshits, S., & Knäuper, B. (2018). Emotional eating and weight regulation: a qualitative study of compensatory behaviors and concerns. Journal of eating disorders, 6, 23.
Ganley, R. M., (1989). Emotion and eating in obesity: a review of the literature. International Journal of Eating Disorders 8, 343/361.
Ge, J. F., Qi, C. C., & Zhou, J. N. (2013). Imbalance of leptin pathway and hypothalamus synaptic plasticity markers are associated with stress-induced depression in rats. Behav. Brain Res. 249, 38–43.
Godet, A., Fortier, A., Bannier, E., Coquery, N., & Val-Laillet, D. (2022). Interactions between emotions and eating behaviors: Main issues, neuroimaging contributions, and innovative preventive or corrective strategies. Reviews in endocrine & metabolic disorders, 23(4), 807–831.
Gold, P. W., & Chrousos, G. P. (2002). Organization of the stress system and its dysregulation in melancholic and atypical depression: high vs low CRH/NE states. Molecular psychiatry, 7(3), 254–275.
Goldsmith, R., Joanisse, D. R., Gallagher, D., Pavlovich, K., Shamoon, E., Leibel, R. L., & Rosenbaum, M. (2010). Effects of experimental weight perturbation on skeletal muscle work efficiency, fuel utilization, and biochemistry in human subjects. American journal of physiology. Regulatory, integrative and comparative physiology, 298(1), R79–R88.
Gluck, M. E., Geliebter, A., Hung, J., & Yahav, E. (2004). Cortisol, Hunger, and Desire to Binge Eat Following a Cold Stress Test in Obese Women with Binge Eating Disorder. Psychosomatic Medicine, 66(6), 876–881.
Gluskin, B. S., & Mickey, B. J. (2016). Genetic variation and dopamine D2 receptor availability: a systematic review and meta-analysis of human in vivo molecular imaging studies. Translational psychiatry, 6(3), e747.
Grajek, M., Krupa-Kotara, K., Białek-Dratwa, A., Staśkiewicz, W., Rozmiarek, M., Misterska, E., & Sas-Nowosielski, K. (2022). Prevalence of Emotional Eating in Groups of Students with Varied Diets and Physical Activity in Poland. Nutrients, 14(16), 3289.
Greeno, G. G., & Wing, R. R. (1994). Stress-induced eating. Psychological Bulletin, 115, 444–464.
Gressier, F., Calati, R., & Serretti, A. (2016). 5-HTTLPR and gender differences in affective disorders: A systematic review. Journal of affective disorders, 190, 193–207.
Grosso, G., Galvano, F. ,Marventano, S., Malaguarnera, M., Bucolo, C., Drago, F., et al. (2014). Omega-3 fatty acids and depression: scientific evidence and biological mechanisms. Oxid. Med. Cell. Longev. 2014, 313570.
Guo, M., Huang, T. Y., Garza, J. C., Chua, S. C., & Lu, X. Y. (2013). Selective deletion of leptin receptors in adult hippocampus induced depression-related behaviors. Int. J. Neuropsychopharmacol. 16, 857–867.
Guo, M., Lu, Y., Garza, J. C., Li, Y., Chua, S. C., Zhang, W., et al. (2012). Forebrain glutamatergic neurons mediate leptin action on depression-like behaviors and synaptic depression. Transl. Psychiatry 2, e83.
Haddock, C. K., & Dill, P. L. (2000). The effects of food on mood and behavior: Implications for the addictions model of obesity and eating disorders. Physiology and Behavior, 70, 333–342.
Haedt-Matt, A. A., Keel, P. K., Racine, S. E., Burt, S. A., Hu, J. Y., Boker, S., Neale, M., & Klump, K. L. (2014). Do emotional eating urges regulate affect? Concurrent and prospective associations and implications for risk models of binge eating. The International journal of eating disorders, 47(8), 874–877.
Hariri, A. R., & Holmes, A. (2006). Genetics of emotional regulation: the role of the serotonin transporter in neural function. Trends in cognitive sciences, 10(4), 182–191.
Heatherton, T. F., Herman, C. P., & Polivy, J. (1991). Effects of physical threat and ego threat on eating behavior. Journal of Personality and Social Psychology, 60(1), 138–143.
Herman, C. P., & Mack, D., (1975). Restrained and unrestrained eating. Journal of Personality 43, 647/660.
Herman, C. P., & Polivy, J. (1975). Anxiety, restraint, and eating behavior. Journal of Abnormal Psychology, 84(6), 666–672.
Herman, C. P., & Polivy, J., (1980). Restrained eating. In: Stunkard, A.B. (Ed.), Obesity. Saunders, Philadelphia.
Herman, C.P., & Polivy, J., (1984). A boundary model for the regulation of eating. In: Stunkard, A.B., Stellar, E. (Eds.), Eating and its disorders. Raven Press, New York, pp. 141/156.
Hryhorczuk, C., Sharma, S., & Fulton, S. E., (2013). Metabolic disturbances connecting obesity and depression. Front. Neurosci. 7:177.
Huether, G., Zhou, D., Schmidt, S., Wiltfang, J., & Ruther, E. (1997). Long-term food restriction down-regulates the density of serotonin transporters in the rat frontal cortex. Biol. Psychiatry 41, 1174–1180.
Jansen, A. (1998). A learning model of binge eating: Cue reactivity and cue exposure. Behaviour Research and Therapy, 36, 257–272.
Jaffe, M., Gullone, E., & Hughes, E. K. (2010). The roles of temperamental dispositions and perceived parenting behaviours in the use of two emotion regulation strategies in late childhood. Journal of Applied Developmental Psychology, 31(1), 47–59.
Kaplan, H. I., & Kaplan, H. S. (1957). The psychosomatic concept of obesity. Journal of Nervous and Mental Disease. 125, 181/201.
Karra, E., O’daly, O. G., Choudhury, A. I., Yousseif, A., Millership, S., Neary, M. T., et al. (2013). A link between FTO, ghrelin, and impaired brain food-cue responsivity. J. Clin. Invest. 123, 3539–3551.
Kirschbaum, C., Pirke, K. M., & Hellhammer, D. H. (1993). The 'Trier Social Stress Test'--a tool for investigating psychobiological stress responses in a laboratory setting. Neuropsychobiology, 28(1-2), 76–81.
Kluge, M.,Schussler, P., Dresler, M., Schmidt, D., Yassouridis, A., Uhr, M., et al. (2011). Effects of ghrelin on psychopathology,sleep and secretion of cortisol and growth hormone in patients with major depression. J. Psychiatry Res. 45, 421–426.
Kluge, M., Schussler, P., Schmid, D., Uhr, M., Kleyer, S., Yassouridis, A., et al. (2009). Ghrelin plasma levels are not altered in major depression. Neuropsychobiology 59, 199–204.
Klump, K. L.(2013). Puberty as a critical risk period for eating disorders: a review of human and animal studies. Horm Behav. 64(2), 399–410.
Klump, K. L., O'Connor, S. M., Hildebrandt, B. A., Keel, P. K., Neale, M., Sisk, C. L., Boker, S., & Alexandra Burt, S. (2016). Differential effects of estrogen and progesterone on genetic and environmental risk for emotional eating in women. Clinical Psychological Science, 4(5), 895–908.
Konttinen, H., Männistö, S., Sarlio-Lähteenkorva, S., Silventoinen, K., & Haukkala, A. (2010). Emotional eating, depressive symptoms and self-reported food consumption. A population-based study. Appetite, 54(3), 473–479.
Köster, E. P., Møller, P., & Mojet, J. (2014). A “misfit theory of spontaneous conscious odor perception (MITSCOP)”: Reflections on the role and function of odor memory in everyday life. Frontiers in Psychology, 5, 64.
Kraus, T., Haack, M., Schuld, A., Hinze-Selch, D., & Pollmächer, T. (2001). Low leptin levels but normal body mass indices in patients with depression or schizophrenia. Neuroendocrinology 73, 243–247.
Lawson, E. A., Miller, K. K., Blum, J. I., Meenaghan, E., Misra, M., Eddy, K. T., et al.(2012). Leptin levels are associated with decreased depressive symptoms in women across the weight spectrum, independent body fat. Clin. Endocrinol. (Oxf.) 76, 520–525.
LeDoux, J. E. (1999). El cerebro emocional. Ed. Planeta.
Leibowitz, S. F., & Alexander, J. T. (1998). Hypothalamic serotonin in control of eating behavior, meal size, and body weight. Biological psychiatry, 44(9), 851–864.
Liu, J., Guo, M., Zhang, D., Cheng, S. Y., Liu, M., Ding, J., et al. (2012). Adiponectin is critical in determining susceptibility to depressive behaviors and has antidepressant-like activity. Proc. Natl. Acad.Sci. U.S.A. 109, 12248–12253.
Liu, J., Perez, S. M., Zhang, W., Lodge, D. J., & Lu, X. Y. (2011). Selective deletion of the leptin receptor dopamine neurons produces anxiogenic-like behavior and increases dopaminergic activity in amygdala. Mol. Psychiatry 16, 1024–1038.
Lombard, C. B. (2000). What is the role of food in preventing depression and improving mood, performance and cognitive function? Med. J. Aust. 173 (Suppl.), S104–S105.
Lowe, M. R., & Fisher, E. B. (1983). Emotional reactivity, emotional eating, and obesity: a naturalistic review. Journal of Behavioral Medicine. 6, 135/149.
Lu, Q., Tao, F., Hou, F., Zhang, Z., & Ren, L. L. (2016). Emotion regulation, emotional eating and the energy-rich dietary pattern. A population-based study in Chinese adolescents. Appetite, 99, 149–156.
Lu, X. Y. (2007).The leptin hypothesis of depression: a potential link between mood disorders and obesity? Curr. Opin. Pharmacol. 7, 648–652.
Lyman, B., (1982). The nutritional values and food group characteristics of food preferred during various emotions. Journal of Psychology 112, 121/127.
Macht, M., (1999). Characteristics of eating in anger, fear, sadness and joy. Appetite 33, 129/139.
Macht, M., (2008). How Emotions Affect Eating: a five-way model. Appetite 50, 1–11.
Macht, M., & Dettmer, D. (2006). Everyday mood and emotions after eating a chocolate bar or an apple. Appetite, 46, 332–336.
Macht, M., & Müller, J. (2007). Immediate effects of chocolate on experimentally induced mood states. Appetite, 49(3), 667–674.
Maffei, M., Halaas, J., Ravussin, E., Pratley, R. E., Lee, G. H., Zhang, Y., et al. (1995). Leptin levels in human and rodent: measurement plasma leptin and ob RNA in obese and weight-reduced subjects. Nat. Med. 1, 1155–1161.
Maniam, J., & Morris, M. J. (2012). The link between stress and feeding behaviour. Neuropharmacology 63, 97–110.
Markus, C. R., Panhuysen, G., Tuiten, A., Koppeschaar, H., Fekkes, D., & Peters, M. L. (1998). Does carbohydrate-rich, protein-poor food prevent a deterioration of mood and cognitive performance of stress-prone subjects when subjected to a stressful task? Appetite, 31, 49–65.
Markus, R., Panhuysen, G., Tuiten, A., & Koppeschaar, H. (2000). Effects of food on cortisol and mood in vulnerable subjects under controllable and uncontrollable stress. Physiology & behavior, 70(3-4), 333–342.
McKenna, R. J., (1972). Some effects of anxiety level and food cues on the eating behavior of obese and normal subjects: a comparison of the Schachterian and psychosomatic conceptions. Journal of Personality and Social Psychology 22, 311/319.
McDowell, D. J., Kim, M., O'Neil, R., & Parke, R. D. (2002). Children's emotional regulation and social competence in middle childhood: The role of maternal and paternal interactive style. Marriage & Family Review, 34(3-4), 345–364.
Miller, A. L. (2005). Epidemiology,etiology, and natural treatment of seasonal affective disorder. Altern. Med. Rev. 10, 5–13.
Morin-Audebrand, L., Mojet, J., Chabanet, C., Issanchou, S., Møller, P., Köster, E. P., et al. (2012). The role of novelty detection in food memory. Acta Psychologica, 139, 233–238.
Morris, W. N., & Reilly, N. P. (1987). Toward The Self-regulation of mood: theory and research. Motiv. Emot. 11, 215–249.
Nederkoorn, C., Smulders, F. T. Y., & Jansen, A. (2000). Cephalic phase responses, craving and food intake in normal subjects. Appetite, 35, 45–55.
Novick, J. S., Stewart, J. W. ,Wisniewski, S. R., Cook, I. A., Manev, R., Nierenberg, A. A., et al. (2005). Clinical and demographic features of atypical depression in outpatients with major depressive disorder: preliminary findings form STAR∗D. J. Clin.Psychiatry 66, 1002–1011.
Oliver, G., & Wardle, J .(1999). Perceived effects of stress on food choice. Physiol. Behav. 66, 511–515.
Ottley, C. (2000). Food and mood. Nurs. Stand. 15, 46–52.
Osman, J. L., & Sobal, J. (2006). Chocolate cravings in American and Spanish individuals: biological and cultural influences. Appetite 47, 290–301.
Ouwens, M. A.,Van Strien, T., & Van Leeuwe, J. F., (2009). Possible pathways between depression, emotional and external eating. Structural Equation model. Appetite 53, 245–248.
Parylak, S. L., Koob, G. F., & Zorrilla, E. P. (2011).The dark side of food addiction. Physiol. Behav. 104, 149–156.
Patel, K. A., & Schlundt, D. G., (2001). Impact of moods and social context on eating behavior. Appetite 36, 111/118.
Pecoraro, N., Reyes, F., Gomez, F., Bhargava, A., & Dallman, M. F. (2004). Chronic stress promotes palatable feeding, which reduces signs of stress: feed-forward and feedback effects of chronic stress. Endocrinology 145, 3754–3762.
Pentikäinen, S., Arvola, A., Karhunen, L., & Pennanen, K. (2018). Easy-going, rational, susceptible and struggling eaters: A segmentation study based on eating behaviour tendencies. Appetite, 120, 212–221.
Pepino, M. Y., Finkbeiner, S., & Mennella, J. A. (2009). Similarities in food cravings and mood states between obese women who smoke tobacco. Obesity (SilverSpring) 17, 1158–1163.
Perello, M., & Zigman, J. M. (2012). The role of ghrelin reward-based eating. Biol. Psychiatry 72, 347–353.
Piazza, P. L. M., & LeMoal, M. (1997). Glucocorticoids as biological substrate of reward: physiological and pathophysiological implications. Brain Res. Rev. 25, 359–372.
Pine, C. J., (1985). Anxiety and eating behavior in obese and nonobese American Indians and White Americans. Journal of Personality and Social Psychology. 49, 774/780.
Pla-Sanjuanelo, J., Ferrer-García, M., Gutiérrez-Maldonado, J., Riva, G., Andreu-Gracia, A., Dakanalis, A., Fernandez-Aranda, F., Forcano, L., Ribas-Sabaté, J., Riesco, N., Rus-Calafell, M., Sánchez, I., & Sánchez-Planell, L. (2015). Identifying specific cues and contexts related to bingeing behavior for the development of effective virtual environments. Appetite, 87, 81–89.
Prasad, C. (1998). Food, mood and health: a neurobiological outlook. Braz. J. Med. Biol. Res. 31, 1517–1527.
Rahme, C., Obeid, S., Sacre, H., Haddad, C., Hallit, R., Salameh, P., & Hallit, S. (2021). Emotional eating among Lebanese adults: scale validation, prevalence and correlates. Eating and weight disorders: EWD, 26(4), 1069–1078.
Raspopow, K., Abizaid, A., Matheson, K., & Anisman, H. (2010). Psychosocial stressor effects on cortisol and ghrelin in emotional and non-emotional eaters: influence of anger and shame. Hormones and behavior, 58(4), 677–684.
Reid, M., & Hammersley, R. (1999). The effects of sucrose and maize oil on subsequent food intake and mood. Br. J. Nutr. 82, 447–455.
Reznick, H., & Balch, P., (1977). The effects of anxiety and response cost manipulations on the eating behavior of obese and normal weight subjects. Addictive Behaviors 2, 219/225.
Rho, S. G., Kim, Y. S., Choi, S. C., & Lee, M. Y. (2014). Sweet food improves chronic stress-induced irritable bowel syndrome-like symptoms in rats. World J. Gastroenterol. 20, 2365–2373.
Rogers, P. J. (1995). Food,mood and appetite. Nutr. Res. Rev. 8, 243–269.
Rossi, S., De Chiara, V.,Musella, A., Mataluni,G., Sacchetti, L., Siracusano, A., et al. (2010). Effects of caffeine on striatal neurotransmission: focus on cannabinoid CB1 receptors. Mol. Nutr. Food Res. 54, 525–531.
Ruderman, A. J., (1983). Obesity, anxiety and food consumption. Addictive Behaviors 8, 235/242.
Ruderman, A. J., (1986). Dietary restraint: a theoretical and empirical review. Psychological Bulletin 99, 247/262.
Sambal, H., Bohon, C., & Weinbach, N. (2021). The effect of mood on food versus non-food interference among females who are high and low on emotional eating. Journal of eating disorders, 9(1), 140.
Sanchez-Villegas, A., & Martinez-Gonzalez, M. A. (2013). Diet, a new target to prevent depression? BMC Med. 11:3.
Schachter, S. (1971). Some extraordinary facts about obese humans and rats. American Psychologist. 26, 129/144.
Schachter, S., Goldman, R., & Gordon, A., (1968). Effects of fear, food deprivation and obesity on eating. Journal of Personality and Social Psychology 10, 90/97.
Schanze, A., Reulbach, U., Scheuchenzuber, M., Groschl, M., Kornhuber, J., & Kraus,T. (2008). Ghrelin and eating disturbances in psychiatric disorders. Neuropsychobiology 57, 126–130.
Schellekens, H., Clarke, G., Jeffery, I. B., Dinan, T. G., & Cryan, J. F. (2012a). Dynamic 5-HT2C receptor editing in a mouse model of obesity. PLoS ONE 7:e32266.
Schmitz, B. A., (1996). The relationship between affect and binge eating. Dissertation Abstracts International: Section B: The Sciences and Engineering 56, 7055.
Schotte, D. E., Cools, J., & McNally, R. J., (1990). Film-induced negative affect triggers overeating in restrained eaters. Journal of Abnormal Psychology 99, 317/320.
Shabbir, F., Patel, A., Mattison, C., Bose, S., Krishnamohan, R., Sweeney, E., et al. (2013). Effect of diet on serotonergic neurotransmission in depression. Neurochem. Int. 62, 324–329.
Sharma, S., & Fulton, S. (2013). Diet-induced obesity promotes depressive-like behaviour that is associated with neural adaptations in brain reward circuitry. Int. J. Obes. (Lond.) 37, 382–389.
Sharma, S., Hryhorczuk, C., & Fulton, S. (2012). Progressive-ratioresponding for palatable high-fat and high-sugar foods mice. J. Vis.Exp. e3754.
Sheppard-Sawyer, C. L., McNally, R. J., & Fischer, J. H., (2000). Film-induced sadness as a trigger for disinhibited eating. International Journal of Eating Disorders 28, 215/220.
Singh M. (2014). Mood, food, and obesity. Frontiers in psychology, 5, 925.
Sleddens, E. F., Kremers, S. P., & Thijs, C. (2008). The children's eating behaviour questionnaire: factorial validity and association with Body Mass Index in Dutch children aged 6-7. The international journal of behavioral nutrition and physical activity, 5, 49.
Slochower, J., Kaplan, S., & Mann, L., (1981). The effects of life stress and weight on mood and eating. Appetite 2, 115/125.
Slochower, J. (1983). Excessive eating: The role of emotions and environment. New York: Human Sciences Press.
Snoek, H. M., Engels, R. C., Janssens, J. M., & van Strien, T. (2007). Parental behaviour and adolescents' emotional eating. Appetite, 49(1), 223–230.
Spinelli S., Masi C., Dinnella C., Zoboli G. P., & Monteleone E. (2014). How does it make you feel? A new approach to measuring emotions in food product experience. Food Qual. Prefer. 37, 109–122.
Spoor, S. T., Bekker, M. H., Van Strien, T., & van Heck, G. L. (2007). Relations between negative affect, coping, and emotional eating. Appetite, 48(3), 368–376.
Stahl, L. A., Begg, D. P., Weisinger, R. S., & Sinclair, A. J. (2008). The role of omega-3 fatty acids in mood disorders. Curr.Opin.Investig.Drugs 9, 57–64.
Souquet, A. M., & Rowland, N. E. (1989). Effect of chronic administration of dexfenfluramine on stress-and palatability-induced food intake in rats. Physiol. Behav. 46, 145–149.
Suzuki, K., Jayasena, C. N., & Bloom, S. R. (2012). Obesity and appetite control. Exp. Diabetes Res. 2012:824305.
Suzuki, K., Simpson, K. A., Minnion, J. S., Shillito, J. C., & Bloom, S. R. (2010). The role of gut hormones and hypothalamus in appetite regulation. Endocr. J. 57, 359–372.
Taylor, G. J., Parker, J. D., Bagby, R. M., & Bourke, M. P. (1996). Relationships between alexithymia and psychological characteristics associated with eating disorders. Journal of psychosomatic research, 41(6), 561–568.
Tomiyama, A. J., Dallman, M. F., & Epel, E. S. (2011). Comfort food is comforting to those most stressed: evidence of the chronic stress response network in high stress women. Psychoneuroendocrinology, 36(10), 1513–1519.
Tryon, M. S., DeCant, R., & Laugero, K. D. (2013). Having your cake and eating it too: a habit of comfort food may link chronic social stress exposure and acute stress-induced cortisol hyporesponsiveness. Physiology & behavior, 114-115, 32–37.
Tschop, M., Smiley, D. L., & Heiman, M. L. (2000). Ghrelin induces adiposity in rodents. Nature 407, 908–913.
Tryon, M. S., Carter, C. S., Decant, R., & Laugero, K. D. (2013). Chronic stress exposure may affect the brain's response to high calorie food cues and predispose to obesogenic eating habits. Physiol.Behav. 120, 233–242.
van Strater, A. C., & Bouvy, P. F. (2006). Omega-3 fatty acids and mood disorders. Am. J. Psychiatry 163:2018.
van Strien T. (2018). Causes of Emotional Eating and Matched Treatment of Obesity. Current diabetes reports, 18(6), 35.
van Strien, T., Herman, C. P., & Verheijden, M. W. (2009). Eating style, overeating, and overweight in a representative Dutch sample. Does external eating play a role?. Appetite, 52(2), 380–387
van Strien, T., Snoek, H. M., van der Zwaluw, C. S., & Engels, R. C. M. E. (2010). Parental control and the dopamine D2 receptor gene ( DRD2 ) interaction on emotional eating in adolescence. Appetite, 54(2), 255–261
van Strien, T., Herman, C. P., & Verheijden, M. W. (2012). Eating style, overeating and weight gain. A prospective 2-year follow-up study in a representative Dutch sample. Appetite, 59(3), 782–789
van Strien, T., van der Zwaluw, C. S., & Engels, R. C. (2010). Emotional eating in adolescents: a gene (SLC6A4/5-HTT) - depressive feelings interaction analysis. Journal of psychiatric research, 44(15), 1035–1042.
van Strien, T., Engels, R. C., Van Leeuwe, J., & Snoek, H. M. (2005). The Stice model of overeating: tests in clinical and non-clinical samples. Appetite, 45(3), 205–213.
van Strien, T., & Ouwens, M. A. (2007). Effects of distress, alexithymia and impulsivity on eating. Eating behaviors, 8(2), 251–257.
van Strien, T., Roelofs, K., & de Weerth, C. (2013). Cortisol reactivity and distress-induced emotional eating. Psychoneuroendocrinology, 38(5), 677–684.
van Strien, T., & Oosterveld, P. (2008). The children's DEBQ for assessment of restrained, emotional, and external eating in 7- to 12-year-old children. The International journal of eating disorders, 41(1), 72–81.
Vandewalle, J., Moens, E., Beyers, W., & Braet, C. (2016). Can we link emotional eating with the emotion regulation skills of adolescents? Psychology & Health, 31(7), 857–872.
Volkow, N. D., Wang, G. J.,Tomasi, D., & Baler, R. D. (2013). Obesity and addiction: neurobiological overlaps. Obes.Rev. 14, 2–18.
Wallin, M. S., & Rissanen, A. M. (1994). Food and mood: relationship between food, serotonin and affective disorders. Acta Psychiatr Scand.Suppl. 377, 36–40.
Ward, A., & Mann, T. (2000). Don't mind if I do: Disinhibited eating under cognitive load. Journal of Personality and Social Psychology, 78, 753–763.
Wei, Q., Lu, X. Y. ,Liu, L., Schafer, G., Shieh, K. R., Burke, S., et al. (2004). Glucocorticoid receptor overexpression in forebrain: a mouse model of increased emotional lability. Proc. Natl. Acad. Sci. U.S.A. 101, 11851–11856.
Williams, J., & Mobarhan, S. (2003). A critical interaction: leptin and ghrelin. Nutr. Rev. 61, 391–393.
Wingenfeld, K., Kuehl, L. K., Boeker, A., Schultebraucks, K., Ritter, K., Hellmann-Regen, J., Otte, C., & Spitzer, C. (2017). Stress reactivity and its effects on subsequent food intake in depressed and healthy women with and without adverse childhood experiences. Psychoneuroendocrinology, 80, 122–130.
Wolz, I., Biehl, S., & Svaldi, J. (2021). Emotional reactivity, suppression of emotions and response inhibition in emotional eaters: A multi-method pilot study. Appetite, 161, 105142.
Wurtman, J. J. (1993). Depression and weight gain: the serotonin connection. J. Affect. Disord. 29, 183–192.
Wurtman, J. J., Wurtman, R. J., Growdon, J. H., Henry, P., Lipscomb, A., & Zeisel, S. H. (1981b). Carbohydrate craving in obese people: Suppression by treatments affecting serotonergic transmission. International Journal of Eating Disorders, 1, 2–15.
Wurtman, R. J., & Wurtman, J. J. (1989). Carbohydrates and depression. Sci. Am. 260, 68–75.
Wurtman, R. J., Hefti, F., & Melamed, E. (1981a). Precursor control of neurotransmitter synthesis. Pharmacological Reviews, 32, 315–335.
Wurtman, R. J., & Wurtman, J. J. (1996). Brain Serotonin, Carbohydrate-craving, obesity and depression. Adv. Exp. Med. Biol. 398, 35–41.
Yau, Y. H., & Potenza, M. N. (2013). Stress and eating behaviors. Minerva Endocrinol. 38, 255–267.
Yehuda, R., Bierer, L. M., Schmeidler, J., Aferiat, D. H., Breslau, I., & Dolan, S. (2000). Low cortisol and risk for PTSD in adult offspring of holocaust survivors. The American journal of psychiatry, 157(8), 1252–1259.
Young, C., & Martin, A. (2003). Omega-3 fatty acids in mood disorders: an overview. Rev. Bras. Psiquiatr. 25, 184–187.
Young, S. N. (2007). Folate and depression–a neglected problem. J. Psychiatry Neurosci. 32, 80–82.
Zagon, A. (2001). Does the vagus nerve mediate the sixth sense? Trends Neurosci. 24, 671–673.
Comentarios
Publicar un comentario